
Wonderful
Life
The Burgess Shale and the
Nature of History
Cat:SCI
Pub:1989 #:0403b
Stephen Jay Gould
00403u/18216r
|
Wonderful LifeThe Burgess Shale and the Nature of History |
Cat:SCI |
Stephen Jay Gould |
||
00403u/18216r |
Title
Wonderful Life
ワンダフル・ライフ
Subtitle
The Burgess Shale and the Nature of History
バージェス頁岩と自然の歴史
Author
Stephen Jay Gould
ステファン. J.ゴールド
Published
1989
1989年
Index
- Preface:
-
- Background for the Burgess Shale:
- Geological time:
- Enigmas:
- Prokaryote:
- Ediacara fauna:
- Tommotian fauna:
- Lagerstatten:
- Walcott's quarry:
- The Age of Arthropod:
- The Burgess Shale Fauna:
- Marrella:
- Yohoia:
- Opabinia:
- Nectocaris:
- Odontogriphus:
- Dinomischus:
- Amiskwia:
- Hallucigenia:
- Canadaspis:
- Aysheaia:
- Odaraia:
- Sidneyia:
- Habelia:
- Leanchoilia:
- Sanctacaris:
- Wiwaxia:
- Anomalocaris:
- In 1985:
- Burgess Shale Fauna:
- The Origin of the Burgess Fauna:
- The first filling of the ecological barrel:
- A directional history of genetic systems:
- Early diversifcation:
- Walcott's Vision:
- Seven Possible Worlds:
Why?
- There is a small quarry in the Canadian Rockies called the Burgess
Shale formed 530 million years ago. It holds the remains of an ancient
sea where dozens of strange creatures lived.
- The current patterns of creatures were not slowly evolved by continuous
proliferation and advance, but set by a pronounced decimation, or
lottery after a rapid initial diversification.
- This book titled "Wonderful Life" deals with the same
theme of the file "It's a Wonderful Life (1946)" of Frank
Capra, whose famous scene was "What the world would be like
without you?"
- "Contingency" is both the watchword and lesson of the
new interpretation of the Burgess Shale.
- カナダロッキー山脈に5.3億年前のバージェス頁岩と呼ばれる採石場がある。そこは古代の海の後でいくつもの奇妙な生物が生きていた。
- 現在の生物のパターンは連続的な増殖と発展による緩い進化の結果ではなく、急激な初期の多様化の後の明確な運不運によって決定した。
- この本の題名である「ワンダフルライフ」は1946年のFrank Capra監督作品1946年の"It's a Wonderful
Life"と同じテーマを扱っている。その有名なシーンで「あなたのいない世界はどうなっただろうか?」
- 「偶然性」はバージェス頁岩の新たな解釈にとっての合い言葉であり教訓である。
Summary
要 約
>Top 0. Preface:
- The beauty of nature lies in detail, while the message, in generality.
- The specific topic is the most precious and important of all fossil
localities - the Burgess Shale of British Colombia.
- Wind back the tape of life to the early days of the Burgess
Shale; let it play again from an identical starting point, and
the chance becomes vanishingly small that anything like human
intelligence would grace the replay.
- The title of this book expressed the duality of our wonder
- at the beauty of the organisms themselves, and the new view
of life that they have inspired.
- Primates are visual animals above all, and anatomical work, in
particular, is as much pictorial as verbal.
- The reconstructions, following a convention in scientific
illustration, rarely show as animal as an observer might have
viewed it on a Cambrian sea bottom - and or two reason. Some
parts are usually made transparent, so the more of
the full anatomy may be visualized; while other parts (usually
the other side of the body) are omitted for the same
reason.
0.序:
- 自然界の美しさは細部に宿り、そのメッセージは一般化の中にある。
- 本話題は、あらゆる化石産地の中で最も貴重で重要なカナダBC州にあるバージェス頁岩である。
- 生命の進化のテープをバージェス頁岩の時代の初期に巻き戻してみる。その時点からすべてをやり直してみると、人類のような知的生物が再び輝く可能性はとてつもなく小さくなる。
- この本のタイトルは二重の意味での驚嘆を表現している。即ち、生命それ自体の美しさとそれがもたらす新たな生命観である。
- 霊長類は何と言っても視覚動物であり、特に解剖学の仕事は言葉と同じように絵で語るものである。
- 科学的なイラストの慣習に従った復元図は、カンブリア紀の海底で見ることができたとしたものとはほとんど別物である。それは、一つには通常ある部分が透明に描かれ解剖的によくわかるようにしてあること、もう一つは同じ理由で体の反対側など一部が省略されていることである。
>Top 1. Replaying Life's Tape:
- The fauna (of the Burgess shale)
was discovered in 1909 by Charles Doolittle Walcott, American paleontologist
and secretary of the Smithsonian Institution.
- This book has three major aims:
- A chronicle of the intense intellectual drama
- It is a statement about the nature of history and the awesome
improbability of human evolution.
- Why such a fundamental program of research has been permitted
to pass to invisibly before the public gaze.
- Some 15-20 Burgess species cannot be allied with any known group,
and should probably be classified as separate phyla.
- Note:
Taxonomy: phylum (division in plants), class, order, family, genus, species,
(subspecies)
- The Burgess Shale includes early representatives of all four major
(Note: 84% of known species) kind of arthropods,
the dominant animals on earth today:
- Trilobites (now extinct)
- Crustaceans (including lobsters, crabs, and shrimp)
- Chelicerates (including spiders and scorpions), and
- Uniramians (including insects).
- The history of life is a story of massive removal followed by
differentiation within a few surviving stocks, not the conventional
tale of steadily increasing excellence, complexity, and diversity.
- If humanity arose just yesterday as a small twig on one branch
of a flourishing tree, then life may not, in any genuine sense,
exist for us or because of us. Perhaps we are only an afterthought,
a kind of cosmic accident, just one bauble on the Christmas
tree of evolution.
1.生命テープの再生:
- バージェス頁岩の動物相は1909年に米国の古生物学者でスミソニアン博物館館長であったCharles
Doolittle Walcottによって発見された。
- 本書には大きく3つの目的がある。
- 激しい知的ドラムの年代記
- 歴史の本質、即ち人間の進化はなかったかもしれないという恐ろしい可能性についての記述
- なぜこれほど重要な研究プログラムが一般の関心を引かずに来たのか。
- バージェス種の15-20種は、どの現生種にも属さない。おそらく別の動物門に分類されるべきである。
- 注):生物の分類:大きな方からphylum門 (植物はdivision), class綱,
order目, family科, genus属, species種, (subspecies亜種)
- バージェス頁岩からは、現在地球上で支配的な(注:既知の種の84%)動物である節足動物門の4つの主要なグループを代表する初期の種が含まれている。
- 三葉虫類 (すでに絶滅)
- 甲殻類 (エビやカニなど)
- 鋏角類 (クモやサソリなど)
- 単枝類 (昆虫など)
- 生命の歴史は、大量の絶滅の後に、生き残った少数の系統の中での分化の物語であって、従来語られたきた優秀さや複雑さや多様性の着実な増大という物語ではない。
- 繁茂する樹木の一本の枝の小枝として、人類が生じたのが昨日だとしたら、いかなる意味でも、生命は人類のためあるいは人類がいるという理由で存在しているのではない。おそらく人類は宇宙の出来事として結果的に生じたものであり、進化のクリスマスツリーにある安物の飾りの一つに過ぎない。
>Top Life's Tapes:
- The sweep of anatomical variety reached a maximum right after the
initial diversification of multicellular animals. The later history
of life proceeded by elimination, not expansion. The current earth
may hold more species than ever before, but most are iterations
upon a few basic anatomical designs.
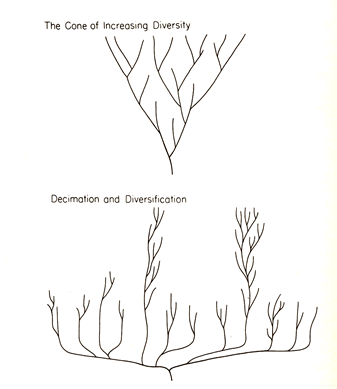 The false but still conventional iconography (upper)
of the cone of increasing diversity, and the revised
model of diversification and decimation (below),
suggested by the proper reconstruction of the Burgess Shale.
The false but still conventional iconography (upper)
of the cone of increasing diversity, and the revised
model of diversification and decimation (below),
suggested by the proper reconstruction of the Burgess Shale.- 'Decimation': (from Latin decimare; to take one in
ten).
The Burgess pattern indicates quite the opposite. Most die and
few are chosen. Decimation has also been used for the taking of
nine in ten; the right metaphor for random elimination of most
lineages.
生命テープ:
- 解剖学的な多様性の広がりが最大に達したのは、多細胞動物が最初の多様化を終えた直後であった。その後の生命の歴史は、拡大ではなく除去によって進展した。現在の地球にはかつてなかったほど多数の種があるように見えるが、その大半は解剖学的には少数の基本的なデザインの反復である。
- (左図) 従来の間違った図式で、多様性が逆円錐状に増加するとされる(上部)とバージェス頁岩の動物群が示唆に基づき修正した多様性と絶滅の復元図 (下部)
- 'Decimation'の意:ラテン語のdecimareより。古代ローマのグループの処刑で、10
人毎に1人をくじで選んで処刑した。バージェス頁岩でのパターンでは全く逆である。ほとんどが絶滅しごく少数が選ばれる。Decimationには10の内9を選ぶという用法もあるので、それならばほとんどの系統がランダムに絶滅するというメタファーとして適切である。
>Top Number of combinations:
- Suppose that ten of a hundred designs will survive and diversify.
If the ten survivors are predictable by superiority of anatomy,
then they will win each time. But if the ten survivors are fortunate
beneficiaries of odd historical contingencies, then each replay
of the tape will yield a different set of survivors and a radically
different history.
- Total number of combinations for 10 items from a pool of
100 yields more than 17 trillion potential outcomes.
組合せの数:
- 100種類のデザインの内10種類が生き残り多様化するとしよう。その10種類が生き残ったのは解剖学的に優れているためだとすれば、何度リプレイしても勝者はいつも同じである。しかしその10種類が生き残ったのは奇妙な歴史的偶然としたら、テープをリプレイするたびに生き残る組み合わせは異なり、その後の歴史も非常に異なったであろう。
- 100種類から10種類を選ぶ組合せの数は17 兆通り以上となる。
>Top Golden mean (Aurea mediocritas):
- An old tradition, dating at least to Aristotle, advise the prudent
person to stake out a position comfortably toward the middle of
the line. But the game of dichotomy has seriously hampered our thinking
about the history of life.
- Each step proceeds for cause, but no finale can be specified
at the start, and none would ever occur a second time in the same
way, because any pathway proceeds through thousands of improbable
stages.
- God dwells among the details, not in the realm of pure generality.
We make out best approach through small curiosities that rivet our
attention - all those pretty pebbles on the shoreline of knowledge.
黄金の中庸:
- アリストテレスまで遡る古い伝統では、賢人は直線の中央辺りの位置で主張すべきという。しかし二分法の駆け引きが生命進化の歴史を巡る我々の思考をひどく阻害してきた。
- 各段階は原因があって進んでいくが、開始時点では最終到達点は特定できず、また同じことが同じやり方では繰り返されない。なぜならどのような道筋も幾千ものありそうもない段階を通って進行していくからである。
- 神は細部に宿るのであって、純然たる一般性の中には宿らない。そのための最善のアプローチは、我々が関心をもつ小さな好奇心、すなわち知識の波打ち際にあるきれいな小石に取り組むことである。
>Top 2. Background for the Burgess Shale:
- The geological time scale:
is divided hierarchically into eras, periods, and epochs. The boundaries
of the largest divisions - the eras - mark the great events.
- The late Cretaceous mass extinction, some 65 million years
ago, sets the boundary between Mesozoic and Cenozoic eras.
- The second boundary, between the Paleozoic and Mesozoic eras,
225 million years ago, records the granddaddy of all extinctions.
- The third and oldest boundary, between Precambrian times and
the Paleozoic era, about 570 million years ago, marks a different
and more puzzling kind of event - "the
Cambrian explosion," or first appearance
of multicellular animals with hard parts in the fossil record.
2.バージェス頁岩の背景: 地質年代区分
- 地質年代のスケールは、大区分から小区分へと順に代・紀・世と階層的に区分される。一番大きな代の境界は最大の出来事境界である古生代と中生代との境は225百万年前で、史上最大規模の大量絶滅があった。
- 三つ目の最古の境は、先カンブリア時代と古生代との境で、約570百万年前であり、他とは異なり謎めいている。それはカンブリア紀の爆発と呼ばれる出来事で、この時に硬い殻をもつ多細胞動物が化石の記録として初めて登場した。
>Top Geological Time:
Era
Period
Epoch
M. yr
代/紀/世
Cenozoic
Quaternary
Holocene
Pleistocene
現世
更新世
Tertiary
Pliocene
Miocene
Oligocene
Eocene
Paleocene
鮮新世
中新世
漸新世
始新世
暁新世
Mesozoic
Cretaceous
65
白亜紀
Jurassic
ジュラ紀
Triassic
三畳紀
Paleozoic
Permian
225
二畳紀
Carbo-
niferous
石炭紀
Devonian
デボン紀
Silurian
シルル紀
Ordovician
オルドビス紀
Cambrian
カンブリア紀
Pre-cambrian
570
先カンブリア時代
>Top Enigmas:
- The Cambrian explosion is a tolerably ancient event, but the earth
is 4.5 billion years old, so multicellular life of modern design
occupies little more than 10% of earthly time; enigmas that obsessed
Darwin and remain central riddles of life's history.
- Schidlowski (1988): Of the two common isotopes of carbon,
C12 and C13, photosynthesis differentially used the lighter
C12 and therefore raised the ratio of isotopes - C12/C13.
- Both stromatolites (mats of sediment trapped and
bound by bacteria and blue-green algae) and actual cells have
been found in the earth's oldest unmetamorphosed sediments,
dating to 3.5 - 3.6 billion years in Africa and Australia.
難題:
- カンブリア紀の爆発も古い時代の出来事だが、地球の年齢は45億年である。従って現代的なデザインを備えた多細胞生物の歴史は、地球の歴史の10%を占めるに過ぎない。これはダーウィンにとっても難題であったばかりか、今でも生命進化の重要な謎となっている。
- Schidlowski (1988)は、炭素の一般的な同位体であるC12とC13の内、光合成は軽い方のC12を特異的に利用し、C12/C13の同位対比率は高くなる。
- ストロマトライト (バクテリアや藍藻に捕らえられ固化した層状堆積物) や実際の細胞が編成されていないアフリカおよび豪州の35-36億年前の最古の堆積岩から見つかっている。
>Top Prokaryote:
- Nearly two-thirds of the entire history of life on earth, all organisms
were single-celled creatures of the simplest, or prokaryotic, design.
- Prokaryotic cells have no organelles - no nucleus, no paired
chromosomes, no mitochondria, no chloroplasts.
- The advent of eukaryotic cells in the fossil record some 1.4
billion years ago.
原核生物:
- 地球上の生命の全歴史の2/3もの間、原核細胞という最も単純なデザインを備えた単細胞生物だけが生息していた。
- 原核細胞は細胞小器官を備えていない。即ち、細胞核も、対をなす染色体も、ミトコンドリアも、葉緑体もない。
- 原核細胞が、化石として登場するのは14億年前である。
>Top Ediacara fauna:
- The Precambrian record does contain one fauna of multicellular animals,
the Ediacara fauna, named for a locality in Australia: Large pancake-flat
soft-bodied creatures.
- The Ediacara is barely Precambrian in age, probably no more
than 700 million years old and perhaps younger.
- The Ediacara animals may represent a failed, independent experiment
in multicellular life, not a set of simpler ancestors for later
creatures with hard parts.
エディアカラ動物群:
- 先カンブリア時代の化石記録には一つの多細胞動物群が含まれている。豪州のエディアカラの丘からエディアカラ動物群と呼ばれる。それらは大きなホットケーキ状のような平坦で体が軟らかい生物。
- エディアカラ動物群はかろうじて先カンブリア時代のものであり7億年前かそれより新しい。
- エディアカラ動物群は、多細胞生物で独立して行われ結局は失敗した実験を示しており、後に登場する硬い殻を備えた生物のより単純な構造の生物群ではなさそう。
>Top Tommotian fauna:
- The first fauna of hard parts, called the Tommotian after a locality
in Russia, contains some creatures with identifiably modern design,
but most of its members are tiny blades, caps, and cups of uncertain
affinity. ("small shelly fauna")
- Instead of Darwin's gradual rise to mounting complexity, the
100 million years from Ediacara to Burgess may have witnessed
three radically different faunas.
トモティ動物群:
- ロシアの地名にちなんでトモティ動物群と呼ばれる硬い殻の最初の動物群が見つかり、現代的なデザインと認められるよいうな生物も含まれているが、大半は小さな水かき、キャップ状やカップ条で、どの部分か関連性がわからない
(有殻微小化石動物群)
- 生物の構造は徐々に複雑になっていったとするダーウィンの説とは異なり、エディアカラからバージェスに至る1億年の間に3種類の全く異なる動物群が登場したを証明しているように見える。
>Top Lagerstatten: ('lode places')
- We designate these faunas of extraordinary completeness and richness
as Lagerstatten. 20% of major groups are known exclusively by: the
Paleozoic Burgess shale, the Devonian Hunsruckschiefer of Germany,
and the Carboniferous Mazon Creek near Chicago.
- Preconditions for the preservation of soft-bodied faunas:
rapid burial of fossils in undisturbed sediment; deposition
in an environment free from the usual agents of immediate destruction
- primarily oxygen and other promoters of decay.
- But the very conditions that promote preservation also decree
that few organisms make their natural home in such places.
- The broad anatomical disparity of the Burgess is an exclusive
feature of the first explosion of multicellular life. No later
Lagerstatten approach the Burgess in breadth of designs for life.
Rather, proceeding forward from the Burgess, we can trace a rapid
stabilization of the decimated survivors.
- The 500 million subsequent years have produced no new phyla,
only twists and turns upon established designs - even if some
variations, like human consciousness, manage to impact the world
in curious ways.
ラーゲルシュテッテン: 鉱脈の場所
- 我々は、特別に完全で豊富な化石動物群をラーゲルシュテッテンと呼ぶ。種王なグループの20%は、古生代のバージェス頁岩 (古生代)
、独のデボン紀フンスリュック頁岩 、およびシカゴ近郊、石炭紀のメゾンクリークだけで発見されている。
- 軟体の動物群が保存される前提条件としては、化石が堆積物中に速やかに埋められ得ること、主として腐敗を促進する酸素などからの破壊から免れるような環境に堆積すること。
- 保存にとって最適な環境は保存すべき生物が本来棲めない環境である。
- バージェス動物群の解剖学的な幅広い異質性は、多細胞生物が最初に爆発的に進化した場合だけに見られる特徴である。バージェスより後の時代のラーゲルシュテッテンでは、生物のデザインの幅の広さにおいてバージェスほどのものはない。むしろバージェス以降では、絶滅の生き残った生物が急速に安定していった跡をたどることができる。
- その後の5億年間は、既製のデザインにひねりや回転が加えられるだけで、新たな動物門は創造されてこなかった。とはいえ人間の意識のような変種が奇妙なやり方で世界に衝撃を与えようとしているが。
 >Top Walcott's quarry:
>Top Walcott's quarry:
- The Burgess Shale occupies one of the most majestic settings - high
in the Canadian Rockies at the eastern border of British Columbia.
Walcott's quarry lies at an elevation of almost 2,400m on the western
slope of the ridge connecting Mt. Field and Mt. Wapta. Turn around
to the west - Emerald Lake below, and the snow-capped President
range beyond, all lit, in late afternoon, by the falling sun. It
resides in Yoho National Park, near the tourist centers of Banff
and Lake Louise.
- At the end of the field season of 1909, when Mrs. Walcott's
horse slid on going down the trail and turned up a slab that at
once attached her husband's attention. Here was a great treasure
- wholly strange Crustacea of Middle Cambrian time.
発掘者Walcottの採掘場:
- バージェス頁岩は最も荘厳な場所にある。そこはカナダBC州の東端に位置するカナディアン・ロッキーの高地にある。Walcottの採掘場はフィールド山とワプタ山をつなぐ稜線の西斜面の高度2,400mにある。西側に目を転じると、眼下にはエメラルド湖が見え、その遠方には雪を頂いたプレジデント山脈が聳え、午後遅くなるとそこに傾いた太陽によってすべてが輝く。そこは多くの観光客が訪れるバンフやルイーズ湖に近いヨーホー国立公園の中に位置している。
- 1909年の野外調査シーズンの終わりに、Walcott夫人が乗っていた馬が山道を下る途中で脚を滑らせて石版をひっくり返し、それは直ちに夫Walcottの注目する所となった。それはカンブリア紀中期の全く目新しい甲殻類の化石というすばらしい宝物だった。
>Top 3. The Age of Arthropods:
- The timing of events becomes jumbled in retrospect, for we arrange
our thoughts in a logical or psychological order that makes sense
to us, not in chronological sequence.
- Freud one remarked that the human mind is like a psychic Rome
in violation of the physical law that two objects can't occupy
the same space a the same time.
- Methodology of research:
Burgess soft parts are not preserved as carbon. By a chemical process
not yet understood, the original carbon was replaced by silicates
of aluminum and calcium, forming a dark reflective layer. The replacement
did not compromise the exquisite preservation of anatomical detail.
- In beginning geology courses, Harry Whittington excelled in
map interpretation and , especially, in drawing block diagrams;
a knack for making three-dimensional structures from two-dimensional
components, and inversely, for depicting solid objects in plane
view. This ability to move from two to three dimensions, and
back again, provided the key for reconstructing the fauna of
the Burgess Shale.
3.節足動物の時代:
- 事件が起きた日時は、振り返ってみるとごちゃ混ぜになっている。我々は自分にとって意味のある論理的または心理的順序で思考を配置するのであって、時間的な順序で配置はしないからである。
- フロイト曰く、人間の心は2つの物体が同時に同一の場所を占められないという物理法則に反する超自然的なローマのようなものである。
- 研究の方法論:
バージェス生物の軟組織は炭素として保存されていない。よくわからない化学作用により、元の炭素はアルミニウムとカルシウムの珪酸塩に置換してしまい、黒色の光沢の層を形成している。その置換が起こっても解剖的な細部まで精緻な保存が損なわれていない。
- 地質学の入門課程で、Harry Whittingtonは地図の解釈、特に地質構造を立体的に描くブロックダイヤグラムに優れていた。即ち二次元の要素から三次元構造を描いたり、逆に立体的な物体を平面に描く才覚があった。この二次元から三次元へ、またその逆の能力はバージェス頁岩の動物群を復元する上でのキーとなった。
-
>Top Classification of Arthropods:
- Don't accept the chauvinistic tradition that labels our era the
age of mammals. This is the age of arthropods. They outnumber
us by any criterion - by species, by individuals, by prospects for
evolutionary continuation. Some 80% of all named animal species
are arthropods, the vast majority insects.
-
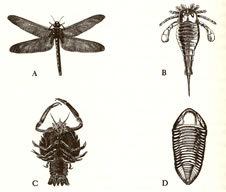 Representative
fossil specimens of the four great groups of arthropods;
Representative
fossil specimens of the four great groups of arthropods;
(A) giant dragonfly; representing Uniramia
(B) eurypterid; Chelicerata
(C) crab; Crustacea
(D) trilobite; Trilobita
- (A) Uniramia: including insects, millipedes,
centipedes, and perhaps also onychophores.
Insects have lost the gill branch; they build their appendages
(antennae, legs, mouth parts) exclusively from leg branches.
- (B) Chelicerata: including spiders, mites,
scorpions, horseshoe crabs, and the extinct eurypterids.
Most modern chelicerates have six uniramous appendages on the
prosoma; The first pair are jawlike and used for grasping. The
second pair are sensory in function. the last four pairs are
leglike. The opisthosomal appendages are also uniramous, but
built from gill branches only.
- (C) Crustacea: including barnacles, decapods
(crabs, lobsters, shrimp)
All crustaceans are distinguished by five pairs of appendages
on the head. The first two pairs, usually called antennae and
antennules, are uniramous. The last three are used in feeding
as mouth parts. Appendages on the trunk often retain the original
biramous form.
- (D) Trilobita: extinct for 225 million years.
The trilobite head bears one pre-oral pair of appendages (antennae)
and three post-oral pairs. Each body segment usually bears a
pair of biramous limbs very little modified from the presumed
ancestral form.
-
節足動物の分類:
- 我々の新生代が哺乳類の時代という熱狂的な伝統を受け入れてはならない。今は節足動物の時代である。種の数、個体数、進化的に存続可能性などどの基準でも哺乳類を凌いでいる。命名されている種の80%は節足動物である、その大部分は昆虫である。
- <左図>節足動物の4大グループを代表する化石
- (A) 単枝類:巨大トンボ (石炭紀)
- (B) 鋏角類:ウミサソリ
- (C) 甲殻類:カニ
- (D) 三葉虫類:三葉虫
- (A) 単枝類:昆虫、ムカデ、ヤスデ、おそらく有爪類。
昆虫類はすべて鰓脚を消失させている。昆虫の付属肢 (触角、肢、口器)は歩脚だけでできている。
- (B) 鋏角類:クモ、ダニ、サソリ、カブトガニ、絶滅したウミサソリ。
現生する鋏角類の大半には頭胸部に六対の単枝型付属肢がある。最初の一対はあごのようになっていて食物をつかむために使われる。二番目の一対は感覚機能がある。残る四対は脚である。但し腹部の付属肢も単枝型であるが、これは鰓脚だけでできている。
- (C) 甲殻類:フジツボ、十脚類 (カニ、エビ)
すべての甲殻類は頭部に五対の付属肢があることで区別できる。最初の二対は単枝型で、触角および小触角と呼ばれる。残り三対は、口器として摂食に使われる。胴体部の付属肢は、原型である二肢型の形をとどめている。
- (D) 三葉虫類:225億年前に絶滅
一対の口前付属肢である触角と三対の口後付属肢がある。それぞれの体節には祖先型からほとんど変化していない一対の二枝型付属肢がある。
-
>Top Metamerism:
- The basic principle of arthropod design is metamerism,
the construction of the body from an extended series of repeated
segments; like a three-part division, such as head, thorax,
and abdomen in insects.
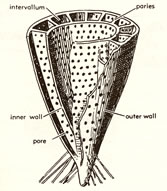
- Exoskeletons:
Stiff, but unmineralized in most groups, thus explaining he rarity
of many arthropods as fossils.
- Biramous (two-branched) limb:
Cross section of an arthropod, showing a pair
of biramous limbs:
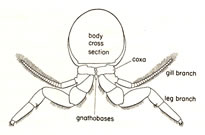 Each appendage consisted of two branches; the inner ramus and the
outer ramus. The outer branch often bear a gill used in respiration
or swimming (gill branch). The inner branch is usually used in locomotion
(leg branch). Specialized arthropods often lose one of the two branches,
retaining the other as a uniramous (one-branched) limb.
Each appendage consisted of two branches; the inner ramus and the
outer ramus. The outer branch often bear a gill used in respiration
or swimming (gill branch). The inner branch is usually used in locomotion
(leg branch). Specialized arthropods often lose one of the two branches,
retaining the other as a uniramous (one-branched) limb.
- Striking phenomenon:
The stereotypy of these patterns is the most striking phenomenon
in modern arthropods. Apparently, evolution settled upon just a
few themes or ground plans for arthropods.
- The story of the Burgess Shale ranks the most amazing in the
history of life: The Burgess shale contains fossils of more
than twenty additional basic arthropod designs.
Why did only four basic designs survive?
-
体節:
- 節足動物のデザインの基本原則は体節である。これは体節構造の繰り返しで体が作られている。昆虫では頭部、胸部、腹部の3つの部分からなる。
- 外骨格:
硬いが、ほとんどの種類で生体鉱物を含まない。このため節足動物の化石は少ない。
- 二枝型付属肢:
<左図>節足動物の断面図、二枝型付属肢を示す:
それぞれの付属肢は、内枝と外枝という2つから成る。外枝には鰓がついていて呼吸あるいは遊泳に使われる (鰓脚)。また内枝は通常は移動のために使われる
(歩脚) 。特殊化した節足動物はしばしばこの二枝の内の一つを失い、付属肢は単枝型となる。
- 驚くべき現象:
これらの定型化したパターンこそが、現生する節足動物の最も注目すべき現象である。進化は節足動物に関してはごく少数の基本設計だけを用意したのである。
- バージェス頁岩をめぐる物語は生命史の中でも最も特筆される。バージェス頁岩には20種類以上の節足動物の基本デザインをもつ化石が発見されている。そしてなぜ4種類の基本デザインだけが生き残ったのだろうか。
>Top 4. The Burgess Shale
Fauna:
-
Marrella: called 'lace crab' by Walcott
- Marrella is a small and elegant
animal. Specimens measure from 2.5 to 19 mm in length. The head
shield is narrow , with two prominent pairs of spines directed
backwards. behind the head, 24 to 26 body segments, each bear
a pair of biramous appendages, composed of a lower walking leg
and an upper branch bearing long and delicate gills. A tiny
button, called a telson, caps the rear end.
- Walcott had called Marrella a trilobite, or at least close
enough to share the anatomical signature of the group.
- Whittington had found that the legs of the body segments are
not established a sequence of head appendages - two pre-oral
and none post-oral - not only unlike the one pre-oral and three
post-oral of trilobites but also completely unknown among arthropods.
- Whittington realized that he would have to abandon Trilobitoidea as an artificial group, a "wastebasket" hiding the
most interesting story of arthropod evolution.
4.バージェス動物群:
マルレラ:
- マルレラは優雅で小さな動物である。体調は2.5-19mm。頭部の背甲は幅が狭く、二対の目立つ長い棘が後方に突き出ている。頭部の後方は24-26対の体節があり、それぞれの体節には一対の二枝型付属肢が生えている。各々の付属肢は、歩脚とその上部に長く繊細な鰓肢で構成されている。
(Walcottはレース蟹と呼んだ) 体節の端は尾節と呼ばれる小さなボタンで終わっている。
- Walcottはマルレラを三葉虫類あるいは少なくとも三葉虫類と解剖的特徴を有するとした。
- Whittingtonは、マルレラの体節から生えてる頭部付属肢の配列が、口前に二対あるだけで口後には一対もないという様式であり、三葉虫類にように口前に一対、口後に三対とは違うだけでなく、節足動物としてもまったく未知のものであることを発見した。
- Whittingtonは、人為的な分類としての三葉虫様綱は、節足動物の最も興味深い進化の歴史を隠してしまうゴミ箱であり、放棄すべきであることに気づいた。
>Top Yohoia:
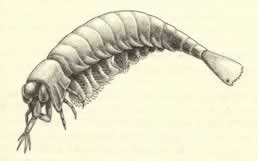
- Yohoia is a very peculiar animal. The head bears three pairs of uniramous
walking legs (the standard trilobite pattern). But the most curious
anomaly of all; the large pair of grasping appendages, composed
of two stout segments at the base and four spines at the tip. This
design is unique among arthropods. Yohoia bears no other appendages
on its head shield - no antennae, no feeding structures. (the so-called
jaws and mouth parts of other arthropods are modified legs) The
first ten body segments behind the head bear lobate appendages fringed
with setae, or hair like extensions.
- Whittington remembers his study of Yohoia as a turning point
in his thinking. He has assimilated Marrella, despite its uniqueness,
under primitive and precursor. But Yohoia forced a different
insight.
ヨホイア:
- ヨホイアは非常に風変わりな動物である。頭部には三対の単肢型の歩脚がある。 (三葉虫の標準的なパターン)しかし非常に奇妙な構造としては、物をつかむことのできる一対の大きな付属肢があり、それは2つのがっしりした節が基部を構成し、その先端には4つの棘がついている。このデザインは節足動物では類のないものである。ヨホイアの頭部背板にはそれ以外の触角や摂食器のような付属肢はない。
(他の節足動物の顎や口器は付属肢が変形したものである。) 後部後方の最初の10節には、剛毛が生えた葉状の付属肢がある。
- Whittingtonは、ヨホイアの研究が転換点になったと語っている。彼は、マルレラは特異な生物であるが、結局は原始的で先駆的なものと見なしていた。しかしヨホイアによって彼は別の考え方を迫られた。
>Top Opabinia:
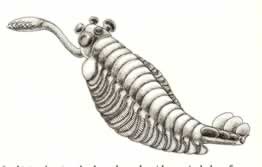
- "When an earlier version of the figure was shown at a meeting
of the Palaeontological Association in Oxford, it was greeted with
loud laughter..." (1975)
Laughter is the most ambiguous of human expressions, for it can
embody two contradictory meanings.
- When we observe according to preset categories, we often cannot
"see" what stares us in the face.
- Actual length: 43-70 mm.
- Opabinia does not have two eyes, but
five! These are arranged as two pairs on short stalk, with a fifth
eye, probably unstalked, mounted on the mid-line.
- The frontal nozzle is attached to the bottom from border of
the head and extends forward. It is a flexible organ, built as
a cylindrical striated tube - literally like the hose of a vacuum
cleaner. Its end is divided longitudinally into two halves, each
with a group of long spines directed inward and forward.
- The main portion of the trunk has 15 segments, each segment
bearing a pair of thin lateral lobes. The lobes overlap, and are
directed downward and outward.
- Each lobe except the first bears on its dorsal surface a paddle
shaped gill attached near the base of the lobe.
- I believe that Whittington's reconstruction of Opabinia in 1975
will stand as one of the great documents in the history of human
knowledge. How many other empirical studies have led directly on
to a fundamentally revised view about the history of life?
オパビニア:
- 「オックスフォード大学での古生物学会において初期の下絵が披露されたとき、それには大笑いが起こった。」(1975) 笑いは相矛盾する2つの意味をもつので、人間の表現してはもっとも曖昧である。
- 我々はプリセットされたカテゴリーに従って観察するとき、我々はしばしば目前にあるものが見えないことがある。
- 体調:43-70mmm
- オパビニアの目は2個ではなくなんと全部で5個ある。その配置は、二対が短い柄の先につき、おそらく柄のない5番目の目は身体の中心線に位置している。
- 前頭部のノズルは頭部下面の最前部に付いて、そこから前方に伸びている。柔軟な器官で、細い溝のついたチューブの管になって、いわば掃除機のホースのようになっている。ノズルの先端は縦方向に二分されており、その内側に長い棘が内向きと前向きに生えている。
- 銅の主要部分には 15の体節があり、各体節の側面には一対の薄い葉状突起が付いている。これらの葉状は互いに重なり合い外側および下向きに突き出ている。
- 先頭部分を除き、個々の葉状突起の背面付け根付近にはオール状の鰓がついている。
- Whittingtonが1975年に発表したオパビニアの復元は、人類の知識獲得の歴史上最大級の成果として語り継がれるものと私は信じる。生命の歴史を巡る観点に根底から修正につながるような経験的研究は他にいくつあっただろうか。
>Top Nectocaris:
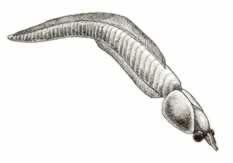
- Nectocaris looks mostly like an arthropod.
The head bears one or two paris of short, forward-projecting, unjointed
(and therefore not arthropod-like) appendages. A pair of large eyes,
probably borne on stalks, lies just behind. The back part of the
head is enclosed by a flattened oval shield, perhaps bivalved. But
the rest of the body evokes no particular hint of arthropod.
- A chimaera: a creature that looks mostly like an arthropod
up front, and mostly like a chordate.
ネクトカリス:
- ネクトカリスは、前部にある首によってほとんど節足動物に見える。頭部からは一対か二対の短い付属肢
(但し間接はないので節足動物とは異なる)が前方に突き出ている。またそのすぐ後ろにおそらく柄の先について一対の大きな目がある。頭部の後半は、楕円形の平たい背板で覆われている。その背板は、おそらく二枚貝のような形状である。しかし体の残りの部分は節足動物を思わせるものは何もない。
- キメラ生物:このように前の部分はほぼ節足動物、後部は脊索動物に見える生物である。
>Top Odontogriphus:
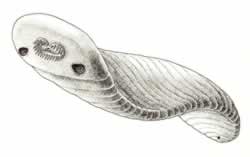
- This highly flattened elongated, oval animal is about 60mm long,
and marked behind its frontal region with a series of fine, transverse
parallel lines, spaced about a mm apart. A pair of "palps"
(probably sensor organs) occupies the corners of the animal's front
end. The more interesting feature, presumably a mouth surrounded
by a feeding apparatus of some kind, lies just forward of the palps,
but right in the mid-line. Conway Morris found some 25 "teeth"
- tiny pointed, conical structures less than half a mm in length.
オドントグリフス:
- これはきわめて扁平で細長いゾウリ型の動物で、体長60mmほどである。全部を除く胴部には約1mm間隔で細い横線が平衡に並んでいる。また一対のヒゲ
(おそらく感覚器官)がこの先端部の隅に位置している。さらに興味深い特徴として、ある種の摂食装置に囲まれた口のような器官が、このヒゲの前方の中心線上にある。Conway
Morrisは、約25本の"歯"があることを発見し、それは長さが0.5mm ほどの先のとがった円錐状のものだった。
>Top Dinomischus:
- Dinomischus is a sessile (fixed and
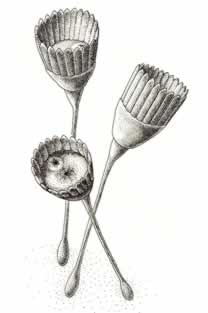 immobile9
creature with radial symmetry, suited to receiving food from all
directions, like many sponges, corals, and stalked crinoids today.
immobile9
creature with radial symmetry, suited to receiving food from all
directions, like many sponges, corals, and stalked crinoids today.
- The goblet, called a calyx, bears on its outer rim a series of
about 20 elongate, parallel-sided blades, called bracts. The upper
surface of the calyx contains both a central and a marginal opening,
presumably mouth and anus by analogy with modern creatures of similar
habits. A U-shaped gut, with an expanded stomach at the base, runs
between the two openings through the interior of the calyx.
ディノミスクス:
- ディノミスクスは、固着性で、すべての方向から来る食物を受け取るのに適した放射状の対称形をしている。現生するものとしては、多くの海綿類、サンゴ類、有柄ウミユリ類とおなじような形をしている。
- 萼 (がく)と呼ばれるのようなゴブレット (足付きグラス) には、その外側の縁に沿って約20個の細長い葉片がずらりと並んでいる。がくの上面には、中心付近と中心をはずれた場所に一個ずつの開口部がある。同様な修正をもつ現生生物との比較から判断すると、これはおそらく口と肛門であろう。がくの内部には、この二つの開口部をつなぐように底部がふくらんだ胃を形成しているU字型の消化管がある。
>Top Amiskwia:
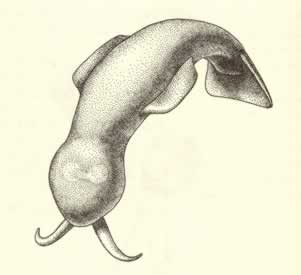
- The head region bears a pair of tentacles, inserted on the front
ventral surface. The trunk sports two fins, unsupported by rays
or any other stiffening device, in the plane of body flattening
- lateral (at the sides) and caudal (forming a tail).
- Amiskwia is probably one of the few Burgess
animals that did not live in the bottom community engulfed by the
mudslide. It was presumably a pelagic (or swimming) organism, living
in open waters above the stagnant basin.
アミスクウィア:
- 頭部には、腹面前部の表面から突き出た一対の触手がある。 胴部には、扁平な体と同一平面をなすように二種類のひれがついている。それらは胴部の両側面にある一対の側ひれと末端部の尾びれであり、いずれのひれにも筋状の支持構造はない。
- アミスクウィアは、泥流に飲み込まれた海底群集の一員ではなかった数少ないバージェス動物の一つであろう。おそらくこの動物は、よどんだ窪地の上の開けた水面で生活する海洋性の遊泳生物だったと思われる。
>Top Hallucigenia:
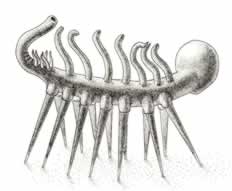
- We need symbols to represent a diversity that we cannot fully carry
in our heads: the stunning disparity and uniqueness of anatomy generated
so early and so quickly in the history of modern multicellular life
- the overwhelming choice among aficionados would surely be Hallucigenia.
How can you describe an animal when you don't even know which side
is up, which end front and which back?
- The largest specimens are about 25mm. Hallucigenia has a bulbous
"head" on one end. This "head" attaches
to a long, narrow, basically cylindrical trunk. Seven pairs
of sharply pointed spines - not jointed, arthropod-like appendages,
but single discrete structures - connect to the sides of the
trunk, near the bottom surface, and extend downward to form
a series of struts.
- Along the dorsal mid-line of the body, directly opposite the
spines, seven tentacles with two-pronged tips extend upward.
The seven tentacles seem to be coordinated with the seven pairs
of spines in an oddly displace but consistent way: the first
tentacle corresponds to no spine below. The last pair of spines
has no corresponding tentacle above.
- A cluster of six much shorter dorsal tentacles (perhaps three
pairs) lies just behind the main row of seven. The posterior
end of the trunk then narrows into a tube and bends upward and
forward.
- The animal couldn't stand there in perpetuity like a stature;
bilaterally symmetrical creatures with heads and tails are almost
always mobile. They concentrate sensory organs up front, and
put heir anuses behind, because they need to know where they
are going and to move away from what they leave behind.
- Perhaps Hallucigenia had no frontal mouth at all. Perhaps
each tentacle gathered food independently, passing the collected
particle down its own personal gullet into the communal gut.
- Perhaps Hallucigenia is not a complete animal, but a complex
appendage of a larger creature, still undiscovered.
ハルキゲニア:
- 我々には、頭では全く想像できないような多様性を実際に示す象徴が必要である。現生する多細胞生物の進化史では、そのごく初期段階に驚くべき早さで、解剖学的デザインの見事な異質性と独自性が生まれてたのである。その実例として熱烈な愛好者が選ぶ生物はといえば間違いなくハルキゲニアであろう。どちらが上でどちらが前でとちらが後かわからない動物をどのように描けば良いのであろうか。
- 体長は最大でも約25mmである。ハルキゲニアには体の一端が球根のような"頭"がある。この"頭"は、細長くて基本的には円筒状の胴にくっついている。鋭く尖った7対の棘
(節足動物の付属肢のような関節はなく、単純で離れた構造である)が胴の両側で底面に近い所に付いていて、一連の支柱のように下方に伸びている。
- 胴体の背面側、即ち棘のある反対側の面の中心線に沿って、先端が二股に分かれた7本の触手が上向きに伸びている。7本の触手は、7対の棘とまさに対応するように規則正しく並んでいる。先頭の一本の触手の下だけは棘がはえていない。最後の一対の棘には上部に対応する触手はない。
- 7本の触手のすぐ後ろの背面には、それよりずっと短い6本 (おそらく3対)の触手がかたまって付いている。そのさらに後ろに続く胴体の端は次第に細く1本の管となり上方さらには前方へと曲がっている。
- この動物は、銅像のようにいつまでもたったままではいかなかった。頭と尾のある左右対称な動物は、必ずと言っていいほど運動能力を備えている。そういう動物は感覚器官を前面に集中させ、肛門は後部に位置している。進行方向のことを知り、後に残していくものから離れる必要があるからである。
- もしかしたらハルキゲニアの先端部には口はなかった可能性がある。それぞれの触手が独立して食物を集め、その集めた食物粒子は独自の食道を通って共同の消化管は送っていたのかも知れない。
- またもしかしたらハルキゲニアは、完全な動物ではなく、まだ未発見のもっと大きな動物の複雑な付属肢かも知れない。
>Top Canadaspis:
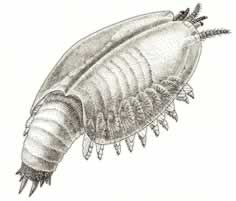
- Canadaspis is the second most common animal in the Burgess Shale. It is large
by Burgess standards (75 mm). This animal is an early malacostracan
(a group of crabs, shrimp, and lobsters) within the Crustacean.
- A head bearing five pairs of appendages, and built of five
segments plus eyes; a thorax (middle section) of eight segments,
and an abdomen (back section) of seven segments plus a telson.
Further, the head appendages are arranged just right, with two
pairs of short, uniramous antennae in front of the mouth, and
three pairs of ventral appendages behind the mouth. The abdominal
segments bear no appendages, but each thoracic segment carries
a par of standard biramous appendages, with an inner leg branch
and a broad outer gill branch.
- Suppose that every Burgess animal were a bizarre denizen of
a lost world. A failed experiment, a washout, a first attempt
totally bypassed by a reconstituted modern fauna, and therefore
offering no clues and no connection to the origin of later life.
But the presence of Canadaspis suggests a different and more
enlightening view. The Burgess fauna does include modern prototypes,
and, in this key respect is an ordinary Cambrian fauna.
カナダスピス:
- カナダスピスは、バージェス頁岩から二番目に多く見つかる動物である。バージェスの標準からいえば大型(75mm)である。この動物は、甲殻類の中の初期の軟甲類
(蟹やエビの仲間) である。
- 頭部は、5対の付属肢があり、5つの体節に目がついたものである。胸部は8つの体節、腹部は7つの体節に1つの尾節という構成である。さらに、頭部の付属肢の配列も適切である。口の前方に2対の短い単肢型の触角があり、口の後方の腹面に3対の付属肢がついている。腹節には付属肢がないが、胸節には一つにつき1対の典型的な二肢型付属肢、つまり内側が歩脚で外側が幅広の鰓脚がある。
- バージェス動物群のすべてが、失われた世界の奇妙な住人たちであるとしよう。その場合、失敗した実験、落伍者、その後再構築される現生動物群に完全に無視された最初の試みとなってしまい、その後の生命の起源についての何の手掛かりも関連性も得られないことになる。ところがカナダピスがいたためにそれとは異なるもっと啓発的な観点が生まれてくる。
バージェス動物群は現生種の祖先を含んでおり、それこそがカンブリア紀によく見られる動物群なのである。
>Top Aysheaia:

- The Onychophora possess a mixture of characters recalling both annelids
and arthropods; many biologists therefore regard this group as one
of the rare connecting forms between two phyla.
- Aysheaia has an annulated, cylindrical
trunk, with ten pairs of annulated limbs attached at the sides
near the lower surface, and pointing down, presumably for use
in locomotion. The anterior end is not separated as a distinct
head. It bears a single pair of appendages. The terminal mouth
is surrounded by six or seven papillae. The head appendages
bear three spinelike branches at their tip, and three additional
spines along the anterior margin.
- The body limbs end in a blunt tip carrying a group of up to
seven tiny, curved claws. Larger spines emerge from the limbs
themselves. These spines are absent on the first pair, point
forward on pairs 2-8, and backward on 9-10.
アユシェアイア:
- 有爪動物は、環形動物と節足動物の両方を思わせる形質を持っている。それ故生物学者はこのグループこそが、2つの動物門を結ぶ希な種類の一つと見なしている。
- アユシェアイアの銅は管節のある円筒状で、その下面に近い側面部から管節のあるおそらく移動用の10対の肢が下向きについている。銅の先端部は頭部として明確に区分されていないが、1対の付属肢が生えている。その付属肢は体側の上の方で水平方向に突き出ている。先端にある口は6本ないし7本の触手に取り囲まれている。頭部の付属肢の先端は、棘状の3本の枝があり、前縁に沿ってさらに3本の棘がある。
- 胴体から生えている肢の先端は途中でちぎれたような終わり方をしており、そこに7本の湾曲した短い爪が生えている。それらの肢の側面には爪よりももっと大きな棘も生えている。これらの棘は一番前の肢にはなく、2-8番目肢では前向き、9-10番目の肢では後ろ向きに生えている。
>Top Odaraia:
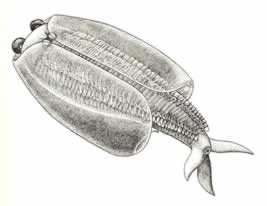
- Odaraia is the largest bivalved arthropod in the
Burgess Shale (150mm).
- The head, with no antennae and only one post-oral pair of
appendages, is unique.
- The trunk, enclosed by the large carapace for more than two-thirds
of its length, contained up to 45 limb-bearing segments. The
limbs, except perhaps for the first two pairs, are typically
bigamous.
- This animal bears a three-pronged tail, with two lateral flukes
and one dorsal projection - a bizarre structure that evokes
images of sharks or whales, rather than lobsters. Nothing similar
exists in any other arthropod. Briggs argued that Odaraia swam
on its back, using its three-pronged tail for stabilization
and steering, and Its carapace as a filtering chamber for capturing
food.
- Briggs had proven once again that he watchword for Burgess
arthropods was "uniquely specialized," not "primitively
simple."
オダライア:
- オダライアは、バージェス頁岩産の二枚貝的な節足動物としては最大 (150mm) 頭部の付属肢としては口の後方に1対の付属肢があるだけで触角はないのはユニークである。
- 全長の2/3以上の胴体は背甲の中に閉じこめられており、最大45個の体節から成る。その体節には肢があるが、最初の2対を除きすべてが典型的な二枝型である。
- この動物は尾びれが3つあり、その内の2つは側面に、1つは背面側に突き出るという奇妙な構造をしている。これはエビの尾びれというより鮫や鯨の尾びれのようである。このような構造は他の節足動物には例がない。Briggsは、オダライアは、3枚の尾を安定装置と舵取り装置として使いながら仰向けで泳いでいたと考え、またその背甲は食物を採るための濾過装置であった。
- Briggsはここでもバージェス産節足動物の合い言葉は「ユニークな特殊化」であって「原始的な単純化」ではないことを証明した。
>Top Sidneyia:
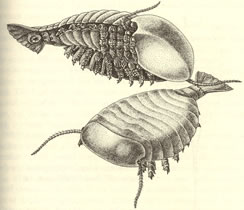
- True chelicerates have six pairs of appendages, and no antennae,
on their head shield. Sidneyia could not
be more different in this crucial respect. Its head bears one pair
of antennae, and no other appendages!
- The first four of nine body segments carry uniramous walking
legs like those of merostomes. But the five posterior segments
bear ordinary biramous appendages, with gill branches and walking
legs. The "tail" piece, formed of three cylindrical
segments and a caudal fan. Bruton interpreted the animal as
a bottom-dwelling carnivore.
シドネユイア:
- 真の鋏角類は、頭部背板に6対の付属肢を備えているが触角はない。 シドネユイアは、この決定的な条件を満たしていない。その頭部は1対の触角がある以外は付属肢はない。
- 9体節の内の最初の4体節には、節口類と同様に単肢型の歩脚がある。一方、後方の5体節には、鰓脚と歩脚からなる通常の二肢型付属肢が付いている。尾に当たる部分は、円筒状の3つの体節と尾びれからなる。Brutonは、この動物は海底に住む肉食者であると解釈した。
>Top Habelia:
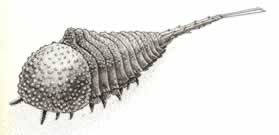 Habelia is blind. The carapace is covered with tubercles. The trunk has
12 segments, with no cylindrical telson. The extended tail spike,
ornamented with barbs and ridges, is unsegmented but has a single
joint about 2/3 of the way back.
Habelia is blind. The carapace is covered with tubercles. The trunk has
12 segments, with no cylindrical telson. The extended tail spike,
ornamented with barbs and ridges, is unsegmented but has a single
joint about 2/3 of the way back.
ハベリア:
- ハベリアには目がない。胴は12の体節からなり、円筒状の尾節はない。 長く伸びた尾の棘はカギ状の突起とうねで飾られ、関節はないが後方2/3ほどの所に継ぎ目が一つだけある。頭部には1対の触角と腹面側に2対の付属肢があるだけである。
>Top Leanchoilia:
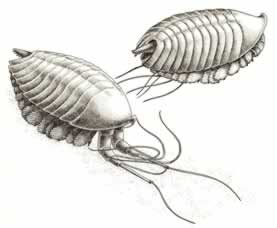
- Top,
in swimming position, with the great appendages folded back and
the whiplike tentacles extending beyond the length of the body;
and bottom, with the great appendages extending forward to aid the
animal in resting on the bottom.
- Leanchoilia does not possess antennae.
It bears 13 pairs of biramous appendages, two at the rear of
the head shield, one on each of the 11 body segments. It bears
such large gill branches that they form a veritable curtain
of overlapping, lamellate lobes, completely covering and extending
beyond the shorter leg branches underneath.
レアンコイリア:
- 上図は遊泳の姿勢で、大付属肢を後方に折りたたんでおり、むち状の触手が体の後方にまで伸びている。下は休憩姿勢。大付属肢を前方に伸ばして海底につけて体を支えている。
- レアンコイリアは、触角がない。13 対の二枝型付属肢があり、その内の2対は頭部背板の後部に、残りは11の体節に1対生えている。その鰓足はとても長く、重なった葉状の薄板が、それより短い歩脚をすっぽり覆い隠しカーテンのように垂れ下がっている。
>Top Sanctacaris:
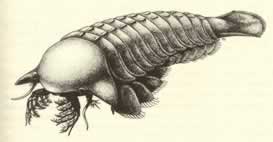
- Sanctacaris has a bulbous head shield, wider than long and extending laterally
as a flat, triangular projection on each side. The body bears eleven
broad segments, the first ten with a pair of biramous appendages.
a wide, flat telson caps the rear end.
- The combination of large lamellate gill branches on the body
appendages and a broad telson well designed for stabilization
and steering indicates that Sanctacaris probably favored swimming
over walking. The striking suite of head appendages identifies
this relatively large arthoropo9d (max 100mm) as a carnivore
specialized for direct pursuit.
サンクタカリス:
- サンクアカリスの頭部背板は、縦より横幅の方が広い球根状で、左右の側面から三角形状の平たい突起が水平に広がっている。胴体は幅広い11体節からなる。最初の10体節には二枝型付属肢が1対ずつ付いている。最後尾は平たくて幅広い尾節となっている。
- 体節に付いている薄板状の大きな鰓脚と、安定性と舵取り機能をもつデザインの尾節をもっていることから、サンクタカリスはおそらく歩行より遊泳が得意だったと見られる。頭部の一対の驚くべき付属肢は、このやや大型の節足動物が、獲物を追いかけて捕らえることに特化した肉食者であることを示している。
>Top Wiwaxia:
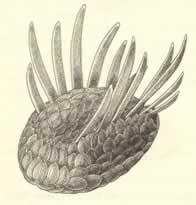
- Wiwaxia is a small creature, shaped as
a flattened oval, about 50mm maximum. Its sclerites grew in two
different styles; flattened scales, ornamented with parallel ridges,
cover most of the body, while two rows of spines emerge from the
top surface, one on each side of the central axis.
- Wiwaxia emerges as a complete, working organism -
a herbivore or omnivore, living on small items of food collected
from the sediment surface as it crawled along the sea floor.
With no setae or appendages and no segmentation, Wiwaxia is neither an arthropod nor an annelid. The jaw displays an
intriguing similarity to the feeding apparatus of mollusks,
called a radula, but nothing else about Wiwaxia even
vaguely resembles a clam, snail, octopus, or any other mollusk
living or dead.
ウィワクシア:
- ウィワクシアは、最大50mmほどの小型生物で、形状は平たい楕円形。その骨片は異なる二種類の形状に生長する。凸凹を作らずに体の全体を覆っている平らな鱗片と、上面から中心軸を挟むように二列に並んで生えている棘である。
- ウィワクシアは植食動物か雑食動物で、海底を徘徊しながら堆積物の表面から小さな食物粒を菜食する生活を送っていたことだろう。剛毛も付属肢も体節もないウィワクシアは、節足動物でも環形動物でもない。そのあごは歯舌と四羽得る軟体動物の摂食装置とよく似ている。しかしそれ以外は二枚貝、巻き貝、イカ、タコなどの現生種や絶滅種も含めどんな軟体動物とも似ていない。
>Top Anomalocaris:
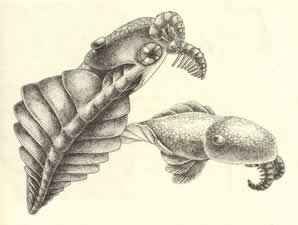
- The
actual chronicle of anomalocaris; culminating in an extraordinary
resolution that brought together bits and pieces of three "phyla"
in a single reconstructed creature, the largest and fiercest of
Cambrian organisms.
- In 1892, J. F. Whiteaves described Anomalocaris (or "odd shrimp") as the headless body of a shrimplike
arthropod.
- The head of Sidneyia?: Walcott joined appendage F
to the head of Sidneyia without direct evidence.
- Walcott identified this fossil as a holothurian, on the basis
of one specimen. He noted a mouth, and thought that it might
be surrounded by a ring of plates.
- Walcott described this peculiar creature as a ring of 32 lobes
around a central opening. Except for radial symmetry, Walcott
found no trace of the defining characters of a jellyfish.
- Now who ever dreamed about a connection between the rear end of
a shrimp, the feeding appendage of Sidneyia, a squashed sea cucumber,
and a jellyfish with a hole in the center?
- One day in 1981, Whittington dug into one end of the creature
and, to his astonishment, found a specimen of Anomalocaris apparently
attached and in place; perhaps this specimen of Anomalocaris
had been accidentally entangled with a large sheet of something
else as the mudslide coalesced?.
- And at last, all the pieces had finally come together. Whittington
and Briggs had reconstructed two separate species of the single
genus Anomalocaris.
アノマロカリス:
- 発見の年代記;最終的には、3つの"動物門"の断片を寄せ集めて、カンブリア紀最大最強の生物が復元されたのである。
- 1892年にJ. F. Whiteavesはアノマロカリス (奇妙なエビの意味)
をエビに似た節足動物の体の後部として記載した。
- シドネユイアの頭?:Walcottは直接の証拠がないのに付属肢Fとしてシドネユアの頭にくっつけた。
- Walcottは、一個の標本をもとにナマコ類として分類した。彼はこの化石の口に注目し、それは骨板で囲まれていたと考えた。
- ここで、エビのしっぽ、シドネユイアの摂食用付属肢、ぺちゃんこのナマコ、中央に穴の開いたクラゲを一つに結びつけることを思いつく者はいるだろうか。
- 1981年のある日、Whittingtonはその化石の一端を掘ってみた。すると驚いたことに、明らかに納まるべき所に納まっているアノマロカリスの標本が姿を現した。この標本も泥流のせいで何らかの大きな薄板の上で偶然合体させられたものかも知れなかった。
- そしてついにすべての部品が集まった。WhittingtonとBriggs はアノマロカリスという単一の属に属する二種の生物を復元した。
>Top In 1985:
- About 600mm in length when extended, the entire animal must
have dwarfed nearly everything else in the Burgess Shale.
- the long oval head bears, on the side and rear portion of
its dorsal surface, a large pair of eyes on short stalks. On
the ventral surface, the pair of feeding appendages attaches
near the front, with the circlet of the mouth behind and in
the mid-line. The mouth may have worked like a nutcracker to
bring prey to the opening, and then crushing its food by constriction.
The teeth may have been attached to the circlet, but they probably
extended from the walls of the gullet.
- Behind the mouth at the ventral surface, the head carries
three pairs of strongly overlapping lobes. The trunk behind
the head is divided into 11 lobes, each triangular in basic
shape, with the apex pointed back in the mid-line. The lobes
are widest at the middle of the trunk. A multilayered structure
of stacked lamellae, presumably a gill, attaches to the top
surface of each lobe.
- Anomalocaris propelled by wavelike motions of the body lobes
in coordinated sequences.
- We do not consider it an arthropod, but the representative
of a hitherto unknown phylum.
1985年:
- 伸ばすと体長は約600mmとなり、カンブリア紀の動物としてはずば抜けた大きさであり、他のほとんどの動物が矮小に見える。
- 楕円形をした頭部の背面上には側面後方に短い柄をもつ1対の大きな目がついている。頭部の腹面には、前縁付近に1対の摂食用付属肢があり、その後方の中心軸上には小環状の口がある。この口はクルミ割りのような機能をがあり、獲物を開口部へ運びかみ砕いていたのだろう。歯は小環に付いていたが、おそらく食道の壁からも伸びていたと思われる。
- 頭部腹面の口の後方には、著しく重なり合った3対の葉状体がついている。胴部は11の葉状体に分かれる個々の葉状体は基本的な形は三角形で、その頂点は中心軸に沿って後方に向いて並んでいる。葉状体は中央部で最も幅が広く、薄板が重なったような多層の構造物が各葉状体の上面にあるが、おそらく鰓であろう。
- アノマロカリスは、胴部の葉状体を連続的に波打つことで推進力を得た。
- 我々はこの動物を節足動物ではなく、現時点では未知の動物門の代表と見なしている。
>Top Burgess Shale
Fauna (Conway Morris and Whittington, 1985):
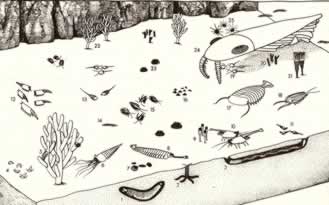
- showing the new interpretation of Anomalocaris
(24), and the great size of this creature compared to the
others.
Note: Opabinia (8), Dinomischus (9), Wiwaxia (23); and the
arthropods Aysheaia (5), Leanchoilia (6), Yohoia (11), Canadaspis
(12), Marrella (15), and Burgessia (19)
バージェス動物群 (Conway
Morris and Whittington, 1985):
- アノマロカリス(24)の新たな解釈と他の生物と比較した場合のずば抜けた大きさ。
注) オパビニア(8)、ディノミスクス(9)、ウィワクシア(23) 、および節足動物のアユシェアイア(5)、レアンコイリア(6)
、ヨホイア(11)、カナダスピス(12)、マルレラ(15)、ブルゲッシア(19)
>Top 5. The Origin of
the Burgess Fauna:
- Three explanations:
Three major kinds of evolutionary explanation are available for
the explosion that led to Burgess disparity.
1) The first filling of the ecological
barrel:
- In conventional Darwinian theory, the organism proposes and the
environment disposes. Organisms provide raw material in the form
of genetic variation expressed in morphological difference. Within
a population at any one time, these differences are small and undirected.
Evolutional change is produced by forced of natural selection arising
from the external environment.
- The Cambrian explosion was the first filling of the ecological
barrel for multicellular life. This was a time of unparalleled opportunity.
Life was radiating onto empty space and could proliferate at logarithmic
rates, like a bacterial cell alone on an agar plate.
- Obvious objections:
- The Permian debacle may have wiped out 95% or more off all
marine species - yet the Burgess phenomenon of explosive disparity
never occurred again.
- 5% may not be a high rate of survivorship, but no mode of
life, no basic ecology, was entirely wiped out by the Permian
debacle.
- If organisms always have the potential for diversification
of this kind - while only the odd ecology of the Lower Cambrian
ever permitted its realization - never, not even once, has a
new phylum arisen since Burgess times.
- I have to believe that organisms as well as environments were
different in Cambrian times, that the explosion and later quiescence
owes as much to a change in organic potential as to an altered
ecological status.
5.バージェス動物群の起源:
- 3つの説明:
バージェスの異質性をもたらした爆発的進化については、主に3つの進化学的な説明が可能である。
1) 生態学的な樽への初の充填:
- 従来のダーウィン理論によれば、生物が提案し、環境が決定する。生物は形態の違いとして現れる遺伝的変異という形で進化の素材を提供する。ある時点でのある個体群内部ではそのような違いは小さくかつ方向性がない。進化的な変化は、外的環境か生じる自然淘汰という圧力によって作られる。
- カンブリア紀の爆発は、多細胞生物にとって生態学的な樽を初めて満たしたのである。当時は、空前絶後の好機に恵まれた時代だった。生命は空っぽの空間へと拡散し、培養地の上に唯一のバクテリア細胞の如く指数関数的に増殖できた。
- 明らかな反論:
- 二畳紀の大量絶滅では、海生生物種の95%以上が一掃されたとされている。にもかかわらず異質性が爆発的に増加するというバージェス的な現象はその後再び起こらなかった。
- 5%は高い生存率とは言えないが、二畳紀の大量絶滅によって、生活様式や基本的な生態系が完全に一掃された訳ではなかった。
- 生物はこのような多様化を遂げる可能性がいつもあるとしたら、下部カンブリア系の特殊な生態学的状況があったがゆえに多様性が実現したとはいうものの、バージェス以降ただの一度も新たな動物門が生じていないのはなぜか。
- カンブリア紀は、環境だけでなく生物も違っていたのではないか。カンブリア紀の爆発とその後の静止状態が、生態学的な状態を変更しただけでなく、生物の能力も同じように変化させたのではないか。
>Top 2) A directional history of genetic
systems:
- In the traditional Darwinian view, morphologies have histories
that constrain their future, but genetic material does not "age."
Differences in rates and patterns of change are responses of an
unchanging material substrate (genes and their actions) to variations
in environment that reset the pressures of natural selection.
- But perhaps genetic systems do "age" in the sense
of becoming "less forgiving of major restructuring".
- Were Cambrian genomes simpler and more flexible?
- Do increase complexity and stereotypy of development from
egg to adult put a brake upon potential changes of great magnitude?
- The discovery of the Precambrian Ediacara fauna, with the
strong possibility that this first multicellular assemblage
may not be ancestral to modern groups, suggests that all Cambrian
animals, despite their disparity of form, may have diverged
not long before from a late Precambrian common ancestor: all
Cambrian animals may have carried a very similar genetic mechanism.
- Life needed the external push of ecological opportunity, but
its ability to respond may have marked a shared genetic heritage,
now dissipated.
2) 遺伝システムとして定方向の歴史:
- 伝統的なダーウィン流の見解によれば、形態変化にはその未来の変化を制限するような過去の歴史がある。但し遺伝物質は「老化」しない。変化の速度とパターンの違いは、不変の物質
(遺伝子とその作用)が、自然淘汰の圧力を改変しようとする環境の差異に対して示す反応の違いである。
- しかしおそらく遺伝システムは、「大規模改造に対しては寛容的でない」という意味において、「老化」するのだろう。
- カンブリア紀のゲノムは、もっと単純で柔軟だったのか。
- 卵から成体への成長過程が複雑かつステレオタイプ化したことで、大規模な変化と遂げる能力にブレーキがかかっているのではないか。
- 先カンブリア紀のエディアカラ動物群が発見され、この最古の多細胞生物群集は現生するグループの祖先ではない可能性が極めて高い。そうだとすると、カンブリア紀のすべての動物は、形態上の異質性にもかかわらず、先カンブリア時代後期の共通祖先から分岐してそれほど経っていないものだった可能性があり、カンブリア紀の動物は、非常によく似た遺伝機構を備えていたであろう。
- 生命は外部から生態学的好機という後押しが必要であった。しかしそれに呼応する能力が、今は霧散しているが当時はまだ共有していた遺伝的な遺産を表出できたのだろう。
>Top 3) Early diversification and later
locking as a property of systems:
- Stu Kauffman has developed a model to demonstrate that the Burgess
pattern of rapid, maximal disparity followed by later decimation
is a general property of systems.
- Changes can be large or small. The opportunity for new body
plans arises with the rarer large jumps. Long jumps are enormously
risky, but yield great reward for rare success.
- How often does a large jump yield a successful outcome? Kauffman
proves that the probability of success is quite high at first,
but drops precipitously and soon reaches an effective zero - just
like the history of life.
- The first few species are place on the landscape at random.
This means that, on average, half the peaks are higher, half lower,
than the initial homes. Therefore, the first long jump has a roughly
50% chance of success. But now the triumphant species stand on
a higher peak - and the percentage of still loftier peaks has
decreased. After a few successful jumps, not many higher peaks
remain unoccupied, and the probability of being able to move at
all drops precipitously. Thereafter, all they can do is hang tough
on their peak or die. Even random extinction leave spaces now
inaccessible to everyone.
- The waiting time to the next higher peak doubles after each
successful jump. If your first success needed only two tries on
average, your tenth will require more than a thousand. Soon you
have effectively no chance of ever getting anywhere better, for
geological time may be long, but it is not infinite.
3) システム特性としての初期の多様化とその後の固定化:
- Stu Kauffmanは、一つのモデルを開発した。それはバージェスに見られる異質性がすばやく最大に達し、その後大量死が続くというパターンは、システムの一般的特性であるという。
- 変化には大きな変化と小さな変化がある。新しい設計図が生み出される機会は、稀に起こる大きな飛躍によってもたらされる。遠くへの飛躍は非常に危険だが、たまに成功した場合の見返りは大きい。
- 大きな飛躍が成功する頻度はどの位かが問題となる。Kauffmanは、成功の確率は最初は極めて高いが、たちまち急降下してすぐにほぼゼロになることを証明した。これは生命進化の歴史と同様である。
- 最初は、少数の種がランダムに地形の上に配置される。これは、平均すれば、移る先の峰がそれまでいた峰より高いか低いかは五分五分の確率ということになる。従って、最初の飛躍で成功する機会は50%である。ところが今や成功した種ほど高い峰に立っているので、それよりさらに高い峰が残っている可能性は少なくなる。最初の数回の飛躍の成功の後では、それ以上に高い未占有の峰はあまり残っていないし、未占有の峰へ移れること自体の可能性が急速に減少する。そうなると後は、自分がいる峰にしがみつくか、死滅するかしかない。もしランダムな絶滅が発生し空き空間ができても、もはや他の生物がそこへ移入することが不可能になっている。
- 飛躍が成功する毎に、次の飛躍が成功するまでの待ち時間は倍々に増加する。飛躍が初めて成功するまでの平均試行回数を2回とすると、10回目の成功を見るまでには1000回以上の試行を重ねなければならない。
>Top 6. Walcott's Vision:
- If Charles Doolittle Walcott had been an ordinary man, his shadow
would not loom so large over the Burgess Shale and his fundamental
error of the shoehorn might merit no more than a footnote. Moreover,
his influence rested squarely upon his deeply conservative and traditional
perspective upon life and morality.
- He was born in 1850 and raised near Utica, New York. Attended
the Utica public schools, but never earned an advance degree.
Joined US Geological Survey in 1879, and raised to Director
in 1894. Then appointed Secretary of the Smithsonian Institution
in 1907. Found the Burgess Shale in 1909. Died in 2/9/1927.
- "Searching for the first form of life", lecture
by Walcott in 1992-94:
Walcott specified the order revealed by paleontology, in a remarkable
passage that embodies the key preconceptions of the shoehorn:
- "In early times the Cephalopoda ruled, later on the Crustacea
came to the fore, then probably Fishes took the lead, but were
speedily outpowered by the Saurians. These Land and Sea Reptiles
then prevailed until Mammalia appeared upon the scene, since
when it doubtless became a struggle for supremacy until Man
was created. Then came the age of Invention, at first of flint
and bone implements,of bows and arrows and fish-hooks, then
of spears and shields, swords and guns, lucifer matches, railways,
electric telegraphs."
- The entire progressionist credo is rolled up into these few words.
Walcott was so committed to progress based on conquest and displacement
that he didn't catch the inaccuracy in his own formulation. Reptiles
did not replace fishes; rather, they represent an oddly modified
group of fishes in a novel terrestrial environment. Fishes have
never been replaced as dominant vertebrates of he oceans.
- For Walcott, the Burgess, as old, had to include a limited
range of simple precursors for later improved descendants. The
modern themes of maximal disparity and decimation by lottery
are more than just unacceptable under such a view of life.
6.ウォルコットのビジョン:
- Charles Doolittle Walcottが並の人間ならば、バージェス頁岩に投げかけられた彼の影響はあれほど巨大ではなかっただろうし、バージェス動物群を既知の分類に「靴べら」で押し込むという彼の根本的な誤りも脚注程度ですんだだろう。彼の影響力は、極めて保守的で伝統的な生命観と倫理観に根ざしている。
- 1850年生まれ。NY州Utica近郊で育つ。Uticaの公立学校に通ったが、それ以上の進学はない。1879年に米国地質調査所に入り、1894年にその所長に就任。1907年にスミソニアン研究所長に就任。1909年にバージェス頁岩を発見。1927年2月9日死去。
- 「最初の生物を探す」Walcottの講演1992-94:
Walcottは、古生物学が解明した順序を特定している。その一節では、「靴べら」を体現した鍵となる先入観がはっきり表明されている。
- 「初期の時代には頭足類が君臨し、続いて甲殻類が頭角を現した。次はどうやら魚類が先頭に躍り出たが、トカゲ類によってすぐに押しのけられた。これら陸と海の爬虫類が幅をきかせたのは哺乳類が舞台に登場するまでだった。哺乳類の登場以降、人間が創造されるまで、間違いなく支配権をかけた闘争の場となった。ついで発明の時代が到来。最初は火打ち石と骨製の道具の発明、次が弓矢と釣り針の発明。あとは槍と楯、剣と銃、黄隣マッチ、鉄道、電報という具合である。」
- 完全な進歩主義者の信条が、上記のいくつかの単語に納まっている。Walcottは、征服と置換を基にした進歩に熱心な余り、自分の公式化の中の間違いに気づかなかった。爬虫類は魚類を置き換えたりはしなかった。むしろ爬虫類は陸上という新しい環境における奇妙に変化した魚類であると言える。魚類は、海洋の支配的な脊椎動物の座を追われてはいない。
- Walcottにとっては、バージェス頁岩は古いのだから、後世の改良された子孫の祖先としての単純な先駆的な生物を含んでいなければならないのである。最大規模の異質性と運任せの悲運な大量死という最近の考えは、このような生命観ではとても認めがたかったのである。
>Top 7. Seven Possible
Worlds:
Evolution of the Eukaryotic Cell:
- 3.5 billion years ago:
Life arose at least 3.5 billion years ago, about as soon as
the earth became cool enough for stability of the chief chemical
components. I suspect that given the composition of early atmospheres
and oceans, life's origin was a chemical necessity.
- The oldest fossils are prokaryotic cells some 3.5 billion
years old; includes stromatolites.
- 1.4 billion years ago:
The first eukaryotic cells (with nucleus and numerous structures
of the cytoplasm) appeared some 1.4 billion years ago. The conventional
argument holds that eukaryotic cells are a prerequisite for
multicellular complexity.
- 570 million years ago:
But multicellular animals did not arise soon after the origin
of eukaryotic cells; they first appeared just before the Cambrian
explosion some 570 million years ago. Hence, more than half
the history of life is a story of prokaryotic cells alone, and
only the last 1/6 of life's time on earth has included multicellular
animals.
(Note: Only a quarter minutes in a
football game!)
- Geological second ago:
Since human intelligence arose just a geological second ago,
we face the stunning fact that the evolution of self-consciousness
required about half of the earth's potential time. Given the
errors and uncertainties, the variations of rates and pathways
in other runs of the tape, what possible confidence we have
in the eventual origin of our distinctive mental abilities?
7.七通りのあり得る世界:
真核細胞の進化:
- 35億年前:
生命は少なくとも35億年前に出現した。それは主な化学物質が安定した状態で存在できる程度に地球が冷えてからすぐのことだった。初期の地球大気と海洋の組合せがあれば、生命の起源は化学的には必然のことではないかと思う。
- 最古の化石は35億年前のストロマトライトなど原核生物のものである。
- 14億年前:
最初の真核細胞 (核と細胞質の多様な器官) は14億年前に登場した。従来の議論では原核細胞は多細胞による複雑化の前提であるとした。
- 5.7億年前:
しかし多細胞動物は原核細胞の起源してからすぐには登場したのではなかった。多細胞動物は5.7億年ほど前のカンブリア紀爆発のほんの少し前に登場している。従って生命の歴史の半分以上が原核細胞しか登場しない物語である。多細胞動物が含まれる期間は、生命が地球に誕生してから今までの時間の後半1/6だけしか多細胞動物は登場しない。
(注:サーカーで言えば後半最後の15分間のみ)
- 地質学的には1秒前:
人類の知性は地質学的には1秒前に登場した。我々は自意識の進化するには地球の許された時間の半分位を必要としたいとう事実に愕然とする。誤差や不確実性、生命テープによっては速度や経路の違いもあろうが、結果として我々の精神的な能力が現れるという確信をどうしたら持てるのだろうか。
>Top Ediacara fauna:

- The
Ediacara fauna has special importance as the only evidence for multicellular
life before the great divide separating the Precambrian and Cambrian.
The Ediacara creatures are only barely Precambrian; they occur in
strata just predating Cambrian and probably do not extend more than
100 million years into the uppermost Precambrian.
- Ediacara design looks like an alternative solution to the
problem of gaining enough surface area as size increases. since
surfaces increase so much more slowly than volumes, and since
animals perform most functions through surfaces. Modern life
followed the path of evolving internal organs (lungs, villi
of the small intestine) to provide the requisite surfaces.
- Ediacara organisms may not be able to evolve internal complexity
and must rely instead on changes in overall form, taking the
shape of threads, ribbons, sheets, or pancakes so that no internal
space lies very far from the outer surface.
エディアカラ動物群:
- エディアカラ動物群は先カンブリア紀とカンブリア紀を隔てる分水嶺の前に存在した多細胞生命の唯一の証拠として特に重要である。エディアカラの生物はかろうじて先カンブリア時代に生きていた。それらはカンブリア紀の直前の地層で、おそらく先カンブリア時代の最後の1億年以前には存在していなかった。
- エディアカラのデザインは、サイズを増加させる一方で十分な表面積を確保するというもう一つの解決法に見える。サイズが増えるとき、体積より面積の方が増加率は小さい。動物はほとんどの機能を体や器官の表面を介して行う。現生生物は、必要な表面積を鯛名期間
(肺や小腸の絨毛) を進化させる道を選んだ。
- エディアカラの生物は複雑な内部構造を進化させられない代わりに全体の形状を変えてより一層、糸状、リボン状、布状、パンケーキ状となることで体腔と外側の体表面を近接させた。
>Top Tommotian creatures:
- The
most characteristic and abundant of all Tommotian creatures, the
archaeocyathids, represent a long-standing problem in classification.
These first reef-forming creatures of the fossil record are simple
in form, usually cone-shaped, with double walls - cup within cup.
Most of the Tommotian oddballs represent unique anatomies that arose
early and disappeared quickly.
トモティ動物群:
- トモティ動物群の中で最も特徴的で豊富なのは古杯類であるが、この分類学上の位置は不明のままである。これら造礁性の生物として初めて化石として登場するこの動物は形状が単純で、その多くは二重の壁を持つ逆円錐形で、杯の中に杯があるような構造をしている。おそらくトモティ動物群の奇妙な形のほとんどは早い時期に登場して早々と消失した解剖学的な特有な形の動物だったかも知れない。
>Top The origin of terrestrial vertebrates:
- The fins of most fishes are entirely unsuited for terrestrial environments.
The few modern fished that scurry across mud flats pull their bodies
along and do not stride with their fins. (walking fish)
- Terrestrial vertebrates could arise because a relatively small
group of fishes, only distantly related to the standard issue,
happen to evolve a radically different type of limb skeleton,
with a strong central axis perpendicular to the body, and numerous
lateral branches radiating from this common focus.
- Wind the tape of life back to the Devonian, the age of fishes.
Would an observer have singles out these uncommon and un characteristic
fishes as precursors to such conspicuous success in such a different
environment?
陸生脊椎動物の起源:
- ほとんどの魚のヒレは陸上環境には全く不向きである。鱈らな泥の上を這い回る現生魚は少数いるが、ヒレでのし歩いていてはいない。(トビハゼ)
- 陸生脊椎動物が生じたのは、比較的少数の魚類で、標準タイプからは遠く隔たったグループが、全く異なるタイプの肢の骨格をたまたま進化させた。それは体から直角に突き出た強力な中心軸をもち、そこから多くの側肢を放射状に伸ばした骨格構造をもつ。
- 生命のテープの魚類の時代であるデボン紀まで巻き戻してみよう。観察者は、このように異なる環境でめざましい成功を収める動物の先駆者として、この変わり者で特徴のない魚を選別できただろうか。
>Top Passing the torch to mammals:
- Mammals evolved at the end of the Triassic, at the same time as
dinosaurs, or just a tad later. Mammals spent their first hundred
million years - 2/3 of their total history - as small creatures
living in the nooks and crannies of a dinosaur's world. Their sixty
million years of success following the demise of dinosaurs has been
something of an afterthought.
- We have no indication of any trend toward mammalian hegemony
during this initial hundred million years. Quite the reverse
- dinosaurs remained in unchallenged possession of all environments
for large-bodied terrestrial creatures. Mammals made no substantial
moves toward domination, larger brains, or even greater size.
- If dinosaurs had not died in this event (extraterrestrial
impact - lucky stars), they would probably still dominate the
domain of large-bodied vertebrates, as they had for so long
with such conspicuous success, and mammals would still be small
creatures in the interstices of their world. This situation
prevailed for a hundred million years, why not for sixty million
more?
哺乳類へ続く希望の光:
- 哺乳類が進化したのは三畳紀の終わりだが、それは恐竜類の進化と同時か少し遅い程度だった。哺乳類は最初の1億年間、その歴史の2/3を恐竜が君臨する世界の片隅や割れ目で生きる小動物として過ごした。恐竜類が滅んだ後の6000万年に及ぶ成功は結果論のようなものだった。
- 哺乳類が登場して最初の1億年の間に、彼らの覇権への傾向が見られたという証拠はない。事実は全く逆で、恐竜類は大型陸上動物にとしてすべての環境を他から脅かされることなく占有し続けた。哺乳類は支配や大きな脳や大型化への移行もなかった。
- もし恐竜類がこの事件(地球外からの幸運な隕石衝突)で死滅していなかったとしたら、おそらくそれらは、それまでのめざましい成功と同様に、それ以降も長きに亘って大型脊椎動物の領域で君臨し、一方、哺乳類は恐竜王国のすきまで生活する小動物のままだっただろう。こういう状態はすでに1億年続いたのであと6000万年続かない理由はない。
>Top The origin of Homo sapiens:
- We view our rise as a kind of global process encompassing
all members of the human lineage, wherever they may have lived.
- Homo sapiens arose as an evolutionary item, a definite
entity, a small and coherent population that split off from
a lineage of ancestors in Africa.
- Asian Homo erectus died without issue and does not
enter our immediate ancestry.
- Neanderthal people were collateral cousins, perhaps
already living in Europe while we emerged in Africa, and also
contributing nothing to our immediate genetic heritage.
- In other words we are an improbable and fragile entity, fortunately
successful after precarious beginnings as a small population
in Africa, not an embodiment of general principles.
人類の起源:
- 我々は、人類の出現は、人類が属する系統の種族が、その住む場所と関係なく世界的規模で進行した出来事だったという見方をしている。
- ホモサピエンスは進化的な細目、明確な実体として登場した。それはアフリカにおいて祖先の系統から分岐した緊密な小集団として登場した。
- アジアのホモエレクトスは子孫を残すことなく死滅したので我々の直接の祖先には入らない。
- ネアンデルタール人は傍系の親類で、我々がアフリカで出現したときにすでにヨーロッパで住んでいた。彼らもまた我々の直接の遺伝的遺産には寄与していない。
- 要するに我々は出現しそうもないもろい実体体だった。アフリカの小集団としておぼつかないスタートの後に幸運にも成功したのであって、一般原理の具体化などではない。
>Top Pikaia:

- Pikaia,
the world's first known chordate. Note the features of our phylum;
the notochord or stiffened rod along the back that evolved into
our spinal column, and the zigzag muscle bands.
- In 1911 Walcott described an attractive species, a laterally
compressed ribbon-shaped creature some 50mm in length. He named
it Pikaia gracilens, to honor nearby Mount Pika. Walcott
confidently placed Pikaia among the polychaete worms.
- Pikaia is not an annelid worm. It is a chordate,
a member of our own phylum; in fact, the first recorded member
of our immediate ancestry.
- The rarity of Pikaia in the Burgess and the absence
of chordates in other Lower Paleozoic Lagestatten, that our
phylum did not rank among the great Cambrian success stories,
and that chordates faced a tenuous future in Burgess times.
- Why do humans exist?
Because Pikaia survived the Burgess decimation. This response
does not cite a single law of nature; it embodies no statement about
predictable evolutionary pathways, no calculation of probabilities
based on general rules of anatomy or ecology. The survival of Pikaia was a contingency of "just history."
ピカイア:
- ピカイアは世界で初めて見つかった脊索動物である。我々の属する動物門の特徴、即ち背面に沿って補強する脊索
(後に我々の脊柱へ進化する) とジグザグ模様の筋帯に注目。
- 1911年にWalcottは、ある魅力的な種として、体調50mm余りのリボン状に扁平した動物を報告している。かれはピカ山にちなんでピカイア・グラキレンスと名付けた。Walcottは自信をもって環形動物門の多毛類と位置づけた。
- ピカイアは環形動物ではない。それは脊索動物であり、我々の直接の祖先として最古の化石となる。
- バージェスではピカイアが稀であること、および他の古生代前期のラーゲルシュテッテンからも脊索動物は発見されていないことから、我々の属する動物門はカンブリア紀には大成功を納めていなかったし、バージェス時代では脊索動物の将来の見通しは暗いものだったろう。
- なぜ人類は存在してるのか:
それはともかくもピカイアがバージェスの悲運な大量死を生き延びたことにある。この答えには自然界の法則は一つも拠り所としていない。そこには予測できる進化の経路に関する言及もなければ、解剖学や生態学の一般法則にもとづいた確率計算もない。ピカイアが生き延びたことは「幸運な歴史」という偶然だった。
Comment
- Many sorts of contingency:
- Contingency is both the watchword and lesson of the new
interpretation of the Burgess Shale.
- Synonym of contingency:
possibility, eventuality, future emergency, uncertainty, something
else....
- Contingency appeared in social life seems more diverse and difficult:
contingency fee, contingency fund, contingency plan, contingency
reserve, contingency table, contingent asset, contingent liability,
contingent damage, contingent beneficiary, contingent interest,
contingent invert, contingent job, ...
- 偶然性いろいろ:
- 偶然性(Contingency)は、バージェス頁岩を巡る新解釈の合い言葉であり教訓である。
- Contingencyの同義語:
possibility 可能性, eventuality 不慮の事件, future emergency 未来の緊急事態, uncertainty
不確実性, something else 何か別のこと....
- 社会生活でのcontingencyはさらに多様で複雑:
contingency fee 成功報酬、 ~ fund 臨時費, ~ plan 予備番組, ~ reserve 偶発損失準備金,
~ table 分割表, contingent asset 偶発資産, ~ liability 偶発債務, ~ damage 偶然損害,
~ beneficiary 次順位受取人, ~ interest 未確定権利, ~ invert 偶発的性倒錯者, ~ job
臨時の仕事 ...
Title |
Wonderful Life |
ワンダフル・ライフ |
|---|---|---|
Subtitle |
The Burgess Shale and the Nature of History | バージェス頁岩と自然の歴史 |
Author |
Stephen Jay Gould | ステファン. J.ゴールド |
Published |
1989 |
1989年 |
Index |
|
|
Why? |
|
|
Summary |
要 約 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top 0. Preface:
|
0.序:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top 1. Replaying Life's Tape:
|
1.生命テープの再生:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Life's Tapes:
|
生命テープ:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Number of combinations:
|
組合せの数:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Golden mean (Aurea mediocritas):
|
黄金の中庸:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top 2. Background for the Burgess Shale:
|
2.バージェス頁岩の背景: 地質年代区分
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Geological Time:
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Enigmas:
|
難題:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Prokaryote:
|
原核生物:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Ediacara fauna:
|
エディアカラ動物群:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Tommotian fauna:
|
トモティ動物群:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Lagerstatten: ('lode places')
|
ラーゲルシュテッテン: 鉱脈の場所
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
発掘者Walcottの採掘場:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top 3. The Age of Arthropods:
|
3.節足動物の時代:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top 4. The Burgess Shale Fauna:
|
4.バージェス動物群:マルレラ:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Yohoia:
|
ヨホイア:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Opabinia:
|
オパビニア:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Nectocaris:
|
ネクトカリス:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Odontogriphus:
|
オドントグリフス:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Dinomischus:
|
ディノミスクス:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Amiskwia:
|
アミスクウィア:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Hallucigenia:
|
ハルキゲニア:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Canadaspis:
|
カナダスピス:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Aysheaia:
|
アユシェアイア:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Odaraia:
|
オダライア:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Sidneyia:
|
シドネユイア:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Habelia:
|
ハベリア:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Leanchoilia:
|
レアンコイリア:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Sanctacaris:
|
サンクタカリス:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Wiwaxia:
|
ウィワクシア:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Anomalocaris:
|
アノマロカリス:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top In 1985:
|
1985年:
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top Burgess Shale
Fauna (Conway Morris and Whittington, 1985):
|
バージェス動物群 (Conway
Morris and Whittington, 1985):
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||
>Top 5. The Origin of the Burgess Fauna:
1) The first filling of the ecological barrel:
|
5.バージェス動物群の起源:
1) 生態学的な樽への初の充填:
|
>Top 2) A directional history of genetic systems:
|
2) 遺伝システムとして定方向の歴史:
|
>Top 3) Early diversification and later locking as a property of systems:
|
3) システム特性としての初期の多様化とその後の固定化:
|
>Top 6. Walcott's Vision:
|
6.ウォルコットのビジョン:
|
>Top 7. Seven Possible Worlds:Evolution of the Eukaryotic Cell:
|
7.七通りのあり得る世界:真核細胞の進化:
|
>Top Ediacara fauna:
|
エディアカラ動物群:
|
>Top Tommotian creatures:
|
トモティ動物群:
|
>Top The origin of terrestrial vertebrates:
|
陸生脊椎動物の起源:
|
>Top Passing the torch to mammals:
|
哺乳類へ続く希望の光:
|
>Top The origin of Homo sapiens:
|
人類の起源:
|
>Top Pikaia:
|
ピカイア:
|
Comment |
|
|